General introduction

Ample experimental data support the notion that investments in fitness trade off with investments in body maintenance. The study of life history regulation in humans does not allow for experimentation but observations of extraordinary phenotypes in specific environments have provided arguments for the notion that human life histories are under similar, genetically encoded control. Virtually all human studies on the topic performed so far used historical cohorts as a source, in part because of the relative ease with which this type of data can be retrieved. Most of these observations regarded people that lived under relatively adverse conditions mimicking the environment in which our population genome has taken shape. These studies have scientific limitations, as other key characteristics of the life histories are scarce whereas access to biomaterials is virtually absent. In contrast, almost all observations of life history regulation in contemporary populations fail from an evolutionary point of view as the subjects under study are exposed to a modern, affluent environment in which fertility is under medical control and mortality patterns have changed dramatically as infection and external causes have disappeared as leading causes of death. It is for these reasons we have started a unique research project in the Upper East Region of Ghana, a remote part of Africa to study determinants of early and late life survival, fertility and fitness under adverse conditions. This scientific endeavour also allowed for collecting more detailed phenotypes and assessment of biomaterials, the ultimate aim being to understand the regulations of human life histories on a biomolecular level. This first chapter provides a very brief introduction to the underlying theories in general and an introduction to the study area more specifically.
Life histories
Evolutionary life history theory argues that ageing is an inevitable consequence of fitness being maximized in a specific environmental niche [1]. In order for the species to survive, investments in fitness, passing genes into subsequent generations go at the cost of investments in maintenance and subsequently leads to a deterioration of the body at older age. This constant trade off between investments in fitness and body maintenance amidst the environmental conditions to which the species is exposed, results in survival probabilities of individuals that are not higher than necessary for reproductive success. Individual life histories are tailored by increasing probabilities of disease and death. Species that have maximized reproductive success at the cost of their individual lifespan are far more likely to survive compared to species that have maximized lifespan but are less reproductively successful.
As early as 1789, some sixty years before Darwin wrote “The origin of species by means of natural selection”[2], Malthus described that all creatures (animals and plants alike) are in constant interaction with the environment to optimise the make up of their own species [3]. Malthus applied this idea to both the animal and plant kingdom. In general, he stated that: ‘many more individuals of a species are born than can possibly survive. As a consequence, there is a constant recurring struggle for existence. The variation of a species that is profitable to itself, will have a better chance of surviving, and will thus naturally be selected. Therefore, any selected variation will tend to propagate its new and modified form.’
In 1801 Lamarck wrote that species are continuously changing over time and argued that individual adaptations to the environment were imprinted and passed into the next generations [4]. This idea was for long time thought to be unrealistic, the force of selection being a superior explanation for the adaptation of the species to its’ environment. Nowadays the ideas of Lamarck are ‘en vogue’ again as epigenetic control of gene expression provides a sufficient biomolecular mechanism how adaptive responses can be passed into the next generation [5].
Some years after Lamarck, in 1813, Wells observed that mulattos and black-skinned people were immune to diseases to which white people were highly susceptible [6]. He postulated that nature selected out races depending on the place they lived. Although Wells did not integrate his observations into theory, amongst others he paved way to the theory of natural selection. Although the theory of evolution and evolutionary selection is still disputed among biologists [7], anthropologists [8,9], theologians and philosophers [10,11], natural selection as the key mechanism for shaping individual life histories cannot be ignored.
Disposable soma
Ageing results from a random accumulation of permanent damage to molecules, cells and tissues [12]. This accumulation primarily results from a continuous, random exposure to amongst others, reactive metabolites, infectious diseases, accidents, famine, and extreme temperatures. The body has the ability to repair some of the damage and to maintain its function. As investments in repair and maintenance go with a cost at fitness, these investments will always be less than is sufficient to prevent the body from deteriorating. The body is thus disposed to allow the genetic lineage to proceed.
Fitness, the success of a genetic lineage, is dependent on fertility per se, and maintenance of the body, in order to survive up to reproductive age. Limited resources have to be divided between body maintenance and fertility. This notion is also described by the r/K-selection theory as proposed by MacArthur and Wilson [13]. The symbols r and K refer to two ends of a continuum, where a compensatory exchange occurs between investment in fertility (r-selection) and in body maintenance (K-selection). Selection for higher investments in maintenance, allowing for longer life spans, is only successful when extrinsic mortality is low enough allowing for reproductive efforts to reduce. For instance, wild mice cannot allow higher investment in maintenance since in their natural environment, exposed to high mortality risks from external causes investments in reproduction are being maximized. Depending on the specific environment, the ‘evolutionary niche’, each species will have optimised investments in body maintenance up to reproductive age and investments in reproductive success to increase the fitness of the species (Figure 1) [14].
The past two decades have brought ample experimental evidence for life history trade-offs within species. Experiments with the fruit fly Drosophila melanogaster proved the general existence of trade-offs between longevity and reproduction, for both females and males [15,16]. A selection regime that favored flies that had retained fertility at later ages resulted in populations with increased life spans, reduced fertility early in life and enhanced resistance to a variety of stresses, suggesting that the mechanisms underlying the increase in lifespan involve greater investments in somatic durability. Direct selection for longevity, by exploiting the dependence of the life span of fruit flies on temperature, also produced long-lived populations with significantly reduced fertility, underpinning a genetic cause for the trade-off [16]. Similar trade-offs have been described for the worm Caenorhabditis elegans. A series of point mutations in the insulin-signaling pathway - regulating metabolism, stress resistance and cell growth - are associated with increases in lifespan of up to 200%, but at the cost of reproductive success [17]. The climax of evidence for the existence of a trade-off between reproduction and longevity, however, is provided by experiments in which germ line precursor cells are removed. As a result, the worm’s lifespan increases markedly [18].
Few would accept that the experimental work in fruit flies and worms would also apply for men, but in recent years several researchers have investigated this trade off between longevity and reproductive success in human populations. Earlier, our group has shown that there is a clear association between longevity and reproductive success using a historical data set from the British aristocracy [19]. We have shown that the number of progeny was small when women died young, increased with an incremental age at death, but decreased again in women who died at an age of 80 years and over. The association was most significant in the pre-1700 period whereas it disappeared in later birth cohorts. This fading of a trade off between longevity and reproductive success suggests that the demographic- and epidemiological transition that started among the aristocracy around 1700 may explain for the disappearance of the negative correlation.
Lycett, Dunbar and Voland [20] examined the same relationship in The Krummhorn region (North-West Germany) in the period 1720 – 1870. They found the association between longevity and reproductive success to be stronger with increasing poverty and successfully argued that their results, in combination with closer inspection of the data among the British aristocracy, suggested that the trade-off between reproduction and longevity is context contingent. The trade off is most apparent under ‘natural conditions’, i.e. when mortality from extrinsic causes is high. These two reports on the relation between longevity and reproduction success have been confirmed in various other populations [21,22,23]. However, some studies have not demonstrated a negative association between fertility and survival. Costa et. al have found no relation between successful aging and reduced reproduction in 88 centenarian women in Calabria, Italy, during the 20th century [24]. Moreover, Le Bourg also found no trade off in a group of French Canadians in the 17th and 18th century [25].
Role of immunity
Here we propose a molecular mechanism that could underlie the tradeoff between reproductive success and longevity, the obvious candidate being immunity [26]. Amongst others, the adverse conditions in our natural habitat necessitate large investments in an adequate immune system to fight abundant infections and therefore reach reproductive age. Earlier we have studied the levels of two major regulatory cytokines — interleukin-10 (IL-10) and tumour necrosis factor (TNF) — in first degree relatives of patients who suffered meningococcal disease, an infection that is widely present in Africa and occasionally surfaces in developed countries [27]. Cytokines are signaling molecules for cell–cell interactions, and include compounds such as TNF that initiate an inflammatory response to fight infection, and regulatory signals such as IL-10 to switch off the inflammatory response and prevent collateral damage after the infection has been overcome. Their activity has been shown to be under tight genetic control and we therefore assumed that families of those patients who had died would have a distinct pattern of cytokine activity. Almost without exception, the level of pro-inflammatory TNF in all of these fatal cases was low, and the level of the anti-inflammatory IL-10 was high [28]. Our interpretation of these data is that subjects with an innate propensity towards anti-inflammatory responses are at an increased risk of death from infection.
In contrast with fighting infection, which requires a strong inflammatory host response,
reproductive success depends on a tolerant immune response. About half of a baby’s
tissue antigens have paternal origin, so at the fetal–maternal interface, immune reactions
must be suppressed to allow pregnancy to proceed. We therefore compared the cytokine profiles of women with impaired fertility, as defined by having at least three
consecutive spontaneous abortions, with the profiles of women of normal fecundity [29]. Reproductive success was associated with a tolerant profile — low TNF and high IL-10 — whereas an inflammatory profile was associated with habitual abortion. The probability of normal fecundity increased up to 16-fold when the women’s cytokine levels were characterized by high anti-inflammatory and low pro-inflammatory profiles.
These data on cytokine profiles help to elucidate two phenomena. First, they can explain why British aristocrats, who lived longer, were less likely to have successful pregnancies. Their innate immune system favored resistance to infection but at the same time prevented pregnancy from proceeding, a trade-off that was even stronger in times when the environmental conditions were relatively poor. Second, it explains why a genotype associated with impaired fertility might have persisted in spite of its obvious disadvantage with regard to evolutionary fitness. Selection for resistance to infection is traded against selection for fertility, resulting in a compromise that is optimal for the fitness of the species in a specific environment.
Despite its protective role in infection, inflammation is potentially harmful; for instance, strong pro-inflammatory responses can cause tissue damage at the site of infection. Although we are programmed to resist infection, in affluent countries the burden of disease has now shifted away from infectious diseases towards chronic diseases that are typically expressed in old age. Accordingly, fatal infections still account for the majority of deaths in less-developed parts of the world, especially at younger ages, but cardiovascular disease has become the leading cause of mortality in ageing populations, accounting for 30% of all deaths worldwide each year. There is clear evidence that inflammation contributes to the development of cardiovascular disease [30]. For example, levels of C-reactive protein, a marker of inflammation, have been associated with coronary artery disease, angina and infarction. Moreover, results from population-based studies have demonstrated that increased levels of markers of inflammation, such as cytokines, adhesion molecules and acute phase reactants, are associated with cardiovascular events. Because our immune system has evolved under the constant attack of pathogens, we are evolutionarily programmed for an inflammatory response to resist infection. In old age, however, the protective effect of this inflammatory response trades off with the increased risk of death from cardiovascular events, thereby reducing life expectancy. Next to cardiovascular disease, inflammation increases the susceptibility and severity of various other chronic diseases.
Aim of the thesis
In this thesis we aim to test the hypothesis that reproductive success trades of with (early) survival in a contemporary population validating the previous findings in historic populations [19,20,23]. For reasons that are outlined above, this test can only be performed in a population that is still exposed to high mortality risk from extrinsic causes and, from an epidemiological point of view, is still in an early stage of transition. After careful analysing three different areas in Africa, we have chosen the remote Garu-Tempane district in the Upper-East Region, Ghana as the research area. This area is one of the least developed areas in Ghana, the socio-economic conditions resembling more of the less developed neighboring countries Burkina Faso and Togo than of southern Ghana. Because of previous research activities in this area [31] and due to the excellent relations with the Ghana Health Service and Regional Healthcare Authorities we were able to access the area quickly.
A short introduction to the research area
The Garu-Tempane district is a remote Upper East Region part of the Republic of Ghana. Over the last centuries it has been an alternating part of Togo, and Gold Coast (former Ghana) [32]. Historically the tribes in this area lived in a constant strife with the ruling Ashanti Kingdom. The Ashanti were intermediary slave traders and recruited their slaves from the tribes of the nowadays Upper Regions [33,34]. In the process of independence (1957) the people living in the eastern part of the Upper East Region (that was under UK-trusteeship) chose to be part of Ghana [32], but the difference in economic development between the south and the north is still enormous. The average per capita income of the southern regions of Ghana in the year 2002 was 2130 dollar [35]. The International Fund for Agricultural Development has estimated the GDP for the Upper East Region at USD 304 per year [36]. Since the Garu-Tempane district is one of the least developed districts in the Upper East Region, the actual GDP of the research area is even lower.
The Garu-Tempane District is densely populated and a total agricultural area (compare: 66 inhabitants per km2 to 43/km2 for the whole Upper East Region). The research area comprises the southern half of this District (see map 2 and 3), measures 375 square kilometer and consists of 36 villages. In total around 24.000 individuals live in the area in 2.300 compounds. The area is inhabited by several tribes, the Bimoba (66%), Kusasi (22%), Mamprusi (4%), Busanga (4%), and Jense (1%) small group of (more nomadic) Fulani (2%) is living in the area as well. Although the Bimoba tribe is by far the largest tribe in the area, they have no ruling power. The area has a semi-Saharan climate with an average temperature of 32 ºC throughout the year and only one rain season (June – August). The entire area has a low level of organization and no civil registries exist.
The vast majority of the population is farmer. The total agricultural process is done by hand labor. Mechanized farming is absent. In the research area some health clinics have been set up recently, but they are not in full service yet. Hospitals and medical services are absent in the area. Vaccination of children was introduced in the early nineties of the previous century. Although not everybody can afford vaccination, at least 50 % of all children under 10 years have been vaccinated at least once. Illiteracy is very high, among adults it is almost complete and among children it is still very high (> 50 %)[37].
Outline
The studies in part A were carried out to analyze the exact stage of epidemiologic transition of the area. The predominant ethnic group within the research area is the Bimoba tribe, living in the southern part of the Garu-Tempane district. Although there is an impressive study of the neighbouring Moba tribe with similar tribe structure and traditions [38], little is known about the Bimoba people [39]. At the time when we started our research, hardly any written information about population and family structure was available. So, to be able to analyse the phenotypic and genetic data, we had to analyse the structure of the research population, the family structure and reproduction patterns first. Chapter 2 is a concise ethnography of the Bimoba. Since the Bimoba tribe is mainly endogamic, consanguinity can become a confounding factor in genetic analysis. Therefore, we analyzed the actual family ties and – possible - inbreeding within the Bimoba tribe. By analyzing the Y-chromosome and the obtained oral pedigree, we were able to draw a genetic pedigree of the population of the FarFar village, a typical village within the area. The results of this study are described in chapter 3. Finally, to establish the actual stage of epidemiologic transition within the region, we analysed the current mortality figures (2002-2005) of the population within the region and compared these estimates with the Gross National Product (GNP) of the area and with all other countries in the world. The results are described in chapter 4.
The second part of the thesis, part B, consisted of testing the trade off between (early) survival and reproductive success from different perspectives. Chapter 5 describes the survival rates of children depending on the size of the kindred as an estimate of fecundity among 2.300 women of 25 years and older. By doing so, we assessed whether the trade off between survival of the children and the reproduction capacity of the mother was present in this population. The production of pro- and anti-inflammatory cytokines among different age groups are compared in Chapter 6. We tested for both the IL-10 and TNF- production and compared the Ghanaian results with the findings in an affluent society, i.e. The Netherlands. We also analyzed known variants of the IL-10 promotor. Together, we tried to find arguments for an immunogenetic explanation for the trade off between (early) survival and reproductive success. The unique DNA collection here gave us the opportunity to test other host response mechanisms as well. In chapter 7 we tested the possible functionality of Caspase-12, an essential protease that plays a crucial role in the inflammatory host response.
Finally, in chapter 8 the results of the entire research project are summarized and discussed in broader perspective.
References.
1. Stearns SC (1992) The Evolution of Life Histories. Oxford University Press, Oxford.
2. Darwin C (1859) The Origin of Species by Means of Natural Selection Or the Preservation of Favoured Races in the Struggle for Life, John Murray, Albemarle Street W, London (used imprint: 1917).
3. Malthus T (1798) An Essay on the Principle of Population, as it Affects the Future Improvement of Society with Remarks on the Speculations of Mr. Godwin, M. Condorcet, and Other Writers, Printed for J Johnson, in St. Paul’s Cathedral-yard London.
4. Lamarck JB (1801) Zoological philosophy: an exposition with regard to the natural history of animals. [transl. from French], (1984) University of Chicago Press, Chicago.
5. Charmantier A, AJ Keyser & DE Promislow (2007) First evidence for heritable variation in cooperative breeding behaviour. Proceedings of the Royal Society 10.1098/rspb.2007.0012 (on line ahead of publication).
6. Wells WC (1813) An Account of a White female, part of whose skin resembles that of a Negro”, Mentioned in C. Darwin (1859) The origin of Species, John Murray Albemarle Street W, London (used imprint: 1917).
7. Duminil J, S Fineschi, A Hampe, P Jordano, D Salvini, et al (2007) Can population genetic structures be predicted from life-history traits? Am Nat. 169(5): 662-72.
8. Claessen H & P Kloos (1978) Evolutie en evolutionisme, Van Gorcum, Assen/Amsterdam.
9. Dobzhansky T (1963) Anthropology and the Natural-Sciences - the Problem of Human-Evolution, Current Anthropology 4:138-139.
10. Dembski W (2002) No Free Lunch. Why specified complexity cannot be purchased without intelligence, Rowman & Littlefield, Lanham, MD
11. Ruse M (2006) The Evolution Creation Struggle, Harvard University Press Cambridge, MA
12. Kirkwood TB (2006) Ageing: Too fast by mistake, Nature 444: 1015 - 1017.
13. Macarthur RH & EO Wilson (1967) The Theory of Island Biography, Princeton University Press, Princeton, New Jersey.
14. Kirkwood TB (1999) Time of our Lifes. The science of Human Aging, Oxford University Press, New York.
15. Sgro CM., G Geddes, K Fowler & L Padrtridge (2000) Selection on age at reproduction in Drosophila melanogaster: female mating frequency as a correlated response, Evolution Int. J. Org. Evolution 54: 2152-2155.
16. Zwaan B, R Bijlsma & RE Hoekstra (1995) Direct Selection on Life-Span in Drosophila-Melanogaster, Evolution 49: 649-659.
17. Arantes-Oliveira N, JR Berman & C Kenyon (2003) Healthy animals with extreme longevity, Science. 302:611.
18. Arantes-Oliveira N, J Apfeld, A Dillin & C Kenyon (2002) Regulation of life span by germ-line stem cells in Caenorhabditis elegans, Science 295: 502-505.
19. Westendorp RGJ, TB Kirkwood (1998) Human longevity at the cost of reproductive success, Nature 396:743-746.
20. Lycett, JE, RI Dunbar & E Voland (2000) Longevity and the costs of reproduction in a historical human population, Proc.Biol.Sci. 267: 31-35.
21. Thomas F, AT Teriokhin, F Renaud, T de Meeus & JF Guegan (2000) Human longevity at the cost of reproductive success: evidence from global data, Journal of Evolutionary Biology 13: 409-414.
22. Smith KR, GP Mineau & LL Bean. (2002) Fertility and post-reproductive longevity, Social Biology 49: 185-205.
23. Doblhammer G (2000) Reproductive history and mortality in later life: A comparative study of England and Wales and Austria, Population Studies 54 169 – 176.
24. Costa MC, F Luzza & R Mattace (2000) Centenarians at no cost of reproductive success, Age and Ageing 29(4): 373-4.
25. Le Bourg E, B Thon, J Legare, B Desjardins & H Charbonneau (1993) Reproductive life of French-Canadians in the 17-18th centuries: a search for a trade-off between early fecundity and longevity, Exp. Gerontol. 28: 217-232.
26. Westendorp RGJ (2004) Are we becoming less disposable? EMBO Rep. 5: 2-6.
27. Westendorp RGJ, A Langermans, TW Huizinga, AH Elouali, CL Verweij, et al. (1997) Genetic influence on cytokine production and fatal meningococcal disease, Lancet 349: 170-173.
28. Dissel JT van, P Langevelde & RGJ Westendorp (1998) Anti-inflammatory cytokine profile and mortality in febrile patients, Lancet 351:950-953.
29. Westendorp RGJ, FM van Dunne, TB Kirkwood, FM Helmerhorst & TWJ Huizinga (2001) Optimising human fertility and survival, Nature medicine 7:873.
30. Libby P, PM Ridker & A Maseri (2002) Inflammation and atherosclerosis, Circulation 105: 1135-1143.
31. Ziem JB, A Olsen, P Magnussen, J Horton, N Spannbrucker et al (2006) Annual mass treatment with albendazole might eliminate human oesophaostomiasis from endemic focus in northern Ghana, Journal of Tropical Medicine & international Health 11(11): 1759-63.
32. Okyere VN (2000) Ghana, a historical Survey, Vinojab Publications, Accra, pp.166-169.
33. Baden-Powell RSS (1898) The Downfall of Prempeh, a diary of life with the native levy in Ashanti, Methuen & Co, London.
34. Meredith M (2006) The State of Africa; A history of fifty years of independence. The Free Press, London, pp. 17 – 29.
35. UN (2002) Community Database World Bank. Online. (assessed 10 may 2007)
36. van Dunne FM, AJ de Craen, FM Helmerhorst, TWJ Huizinga, RGJ Westendorp (2006) Interleukin-10 promoter polymorphisms in male and female fertility and fecundity, Genes Immun 7(8): 688-92.
37. Ghana Statistical Services (2004) Demographic and Health Survey 2003, Noguchi Institute for Medical Research, Accra.
38. Zwernermann J (1998) Studien zur Kultur der Moba (Nord-Togo), Rűdiger Köppe Verlag, Köln.
39. Assimeng M (1990) Bimoba Sociological Study, University of Ghana, Legon.
Legend to the Figures
1. Schematic diagram of the trade off between investments in maintenance (projected as life years) and fitness (projected as number of progeny). (adapted from Kirkwood, 1999).
2. Map of Ghana.
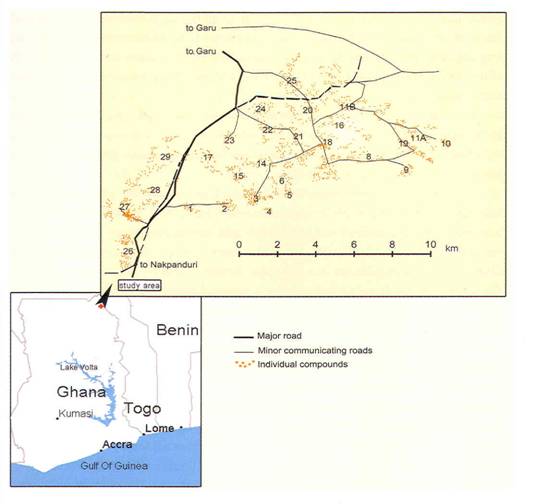
3. Map of Research Area by means of GPS projection (the villages are numbered for research reasons only).
4. Map of a part of Village 3 (FarFar) by means of GPS-projection. (each compound has an individual research number).
Figure 1
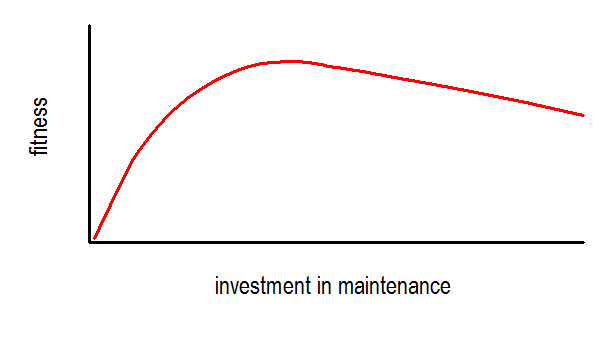
Figure 2